

Evolución de la primera articulación tarsometatarsiana. Un viaje de ida y vuelta
The evolving tarso-metatarsal joint. A go & back travel
Introducción
El objetivo de esta publicación es mostrar los principales cambios morfológicos del cuneiforme medial (CM) y de la primera articulación tarsometatarsiana (1.ª ATMT) a través de los tiempos hasta llegar al ser humano. Estos cambios fueron necesarios para ayudar a la adaptación del pie a las condiciones medioambientales, para que este fuera lo más funcional posible.
Hace unos 350 millones de años (MA), a finales del periodo Devónico, en un periodo de gran sequía, un grupo de peces primitivos colonizaron la interfase agua-tierra firme y empezaron a reptar con la ayuda de sus cuatro aletas. Las anteriores fueron las precursoras de las extremidades anteriores de los cuadrúpedos y de las alas de las aves y las posteriores de las extremidades inferiores, de las que su porción más distal se convertiría en el pie. El más conocido de estos peces es el Eusthenopteron, muy parecido morfológica y taxonómicamente al actual celacanto(1), por lo que algunos autores consideran a este pez como un “fósil viviente”.
Juntamente con la presencia cada vez mayor de aguas poco profundas(2,3), aparecieron los primeros anfibios y, con ello, la deambulación tetrápoda. En los anfibios primitivos, la zona media del pie se componía de una serie de núcleos óseos llamados os tarsale, entre 3 y 5 elementos, y otro núcleo óseo denominado os pretarsale. Estas pequeñas formaciones osteocartilaginosas, con el devenir de los tiempos, se fusionarían para formar los huesos del retropié y del mediotarso(4), donde se incluyen las articulaciones mediotarsiana y tarsometatarsiana.
Durante el periodo Triásico inferior (hacia los 230 MA) se produce la transformación de un pie primitivo (“pie reptil”) en otro moderno (“pie mamífero”). Ello se pudo objetivar a través de los cambios morfológicos vistos en un tipo especial de reptiles-mamiferoides, los Cynodontia(5) (Figura 1).
rpt.3502.fs2104011-figura1.png
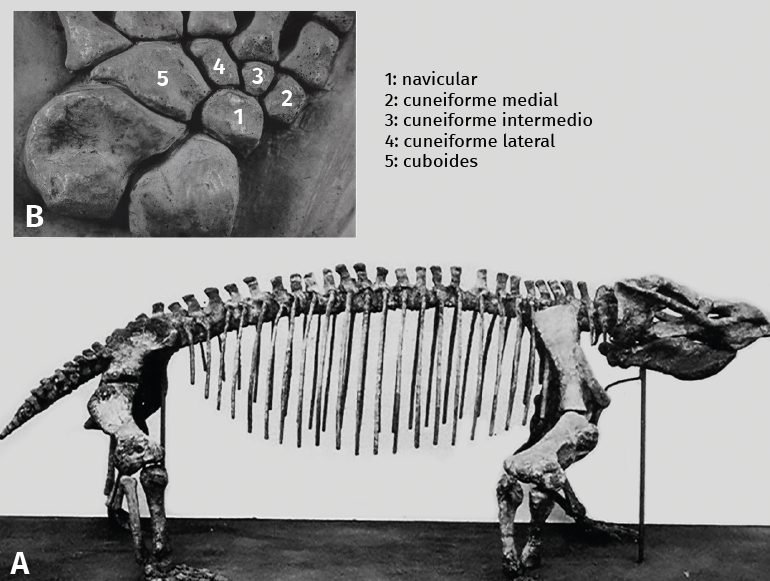
Figura 1. A: reptil mamiferoide avanzado (Aulacoephalodon peavoti/therapsido); B: detalle del mediotarso de un reptil-mamiferoide avanzado (Bauria cynops/Cynodontia). El radio de la derecha es el primer radio; los 3 huesos que coinciden con el 1.º, 2.º y 3.er radios son los 3 cuneiformes.
Poco después, a finales del Triásico/principios del Jurásico (215 MA), aparecieron los primeros mamíferos no placentados (cuyo género más representativo es Eozostrodon) y fue precisamente en este nivel evolutivo donde los diferentes os tarsale se convertirían en los 3 cuneiformes, mientras que el os pretarsale se convertiría en el cuboides(6).
Podemos pues datar en torno a los 215 MA la aparición del CM y de la 1.ª ATMT como entidad morfológica definida.
Pero no todos los radios eran exactamente iguales. Analizando en los restos óseos del pie en estos primitivos animales, se pueden atisbar ligeros cambios en el primer radio (el más medial y anterior durante la deambulación), que van unidos a una ligera divergencia con los otros 4 radios. Este cambio ha podido ser estudiado, principalmente, a través de las diferencias morfológicas de la 1.ª ATMT, diferente a la que tienen los otros radios “externos”. Dicha articulación adopta una forma inicial en silla de montar.
Hace 60 MA, durante el Paleoceno, aparecen los primates arcaicos, cuyo género más estudiado es el Plesiadapis. Sus extremidades inferiores acaban en pies con garras y tanto el hallux como el resto del pie difieren morfológicamente muy poco de otros animales similares como roedores o mustélidos(7).
Hace unos 53 MA, durante el Eoceno, aparecen los primeros primates modernos. Este periodo geológico se caracterizó por una gran proliferación de bosques y de las selvas a causa de un incremento acusado de la temperatura media y una pluviometría elevada. Dicha circunstancia geoclimática hizo que los primates, en este caso los Prosimii extintos, estuvieran adaptados perfectamente a la vida arbórea. Para ello, tanto el hallux como el pollex se convierten en útiles aprehensores a través de una capacidad de divergencia y rotación no vista hasta ese momento en el registro fósil (Figura 2). Esta capacidad de divergencia asociada a la aparición de los dermatoglifos cutáneos palmares y plantares, conjuntamente con la sustitución de las garras por uñas, generaron una gran diversidad de movimientos, la cual se vio incrementada por la diferente inserción de algunos de sus músculos y los cambios morfológicos articulares, principalmente en el primer radio. Los géneros más estudiados por lo que respecta al pie son los géneros Adapis en Eurasia, cuyo tren inferior incluido el pie ha sido estudiado a partir de un ejemplar del yacimiento paleontológico de Messel en Alemania, y Smilodectes en América. Tanto es así, que en estos primates la mayor congruencia articular de la 1.ª ATMT se alcanza en la posición de aprehensión (grasping foot)(8) y en ello influirá de manera determinante una nueva y amplia inserción del tendón del peroneus longus en el CM(9).
rpt.3502.fs2104011-figura2.png

Figura 2. Prosimio. A: Nycticebus concuang; B: gran separación entre el primer radio y los radios laterales.
Hacia los 35 MA, durante el Oligoceno, de nuevo un cambio climático drástico tuvo lugar en nuestro planeta (bajada de la temperatura media de más de 20° y escasez de lluvias en comparación con el periodo anterior, el Eoceno). Estas circunstancias determinarían la aparición de los primates antropoides, capaces de desplazarse en suelo firme. Sin embargo, no perdieron enteramente su capacidad de trepar a los árboles (Figura 3).
rpt.3502.fs2104011-figura3.png
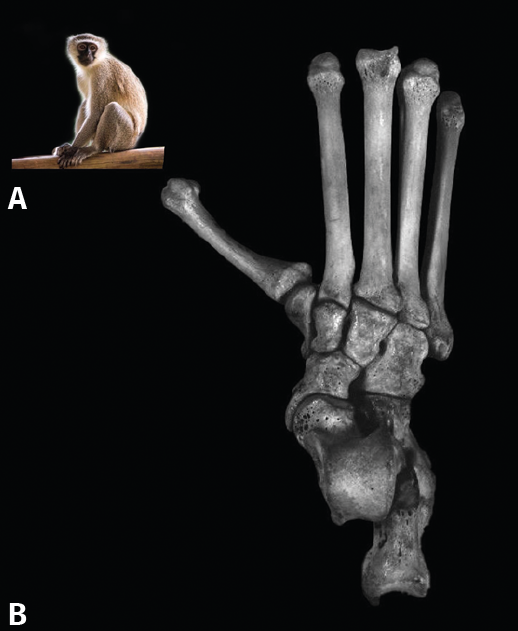
Figura 3. Simio antropoide. A: Cercopithecus aethiops; B: obsérvese la oblicuidad de la primera articulación tarsometatarsiana.
Durante el Mioceno (23 MA), los primates antropoides compensaron la escasa capacidad de abducción del hallux mediante una mayor supinación del segundo metatarsiano.
Todos los datos genéticos indican que a finales del Mioceno (11,6 y 7,2 MA) surgió en algún lugar de África la familia Hominoidea(10). Los géneros actuales de esta familia, enteramente parecidos morfológicamente a otros extintos, muestran entre ellos diferencias morfológicas evidentes en la 1.ª ATMT. Los más primitivos y alejados filogenéticamente del hombre moderno son los Hylobatidae (gibones), los cuales presentan una 1.ª ATMT con un eje vertical, así como la presencia constante de un os pre-hallux en el cual se inserta un fascículo del músculo tibial anterior. Por el contrario, en los Pongidae, más próximos a nosotros (orangután, gorila y chimpancé), la inserción del tendón peroneus longus, que en el hombre está en el CM, se inserta en la base del primer metatarsiano, ayudando a aducir el hallux(11). También son notables las diferencias morfológicas en la 1.ª ATMT. Dicha articulación presenta una doble curvatura, siendo convexa en la parte dorsal y cóncava en la plantar(12).
Datado a finales del Plioceno (3,67 MA), se descubrió en la región sudafricana un individuo que se ha atribuido a Australopithecus prometheus. Aparte de la importancia evolutiva intrínseca, este esqueleto es notable, puesto que se conserva un pie casi completo, StW 57 “Little foot”(13). La 1.ª ATMT es ligeramente cóncavo-convexa, de lo cual se deduce una notable capacidad arborícola. Por el contrario, la morfología de la zona distal del primer metatarsiano es muy parecida al Homo habilis OH 8(14), mucho más moderno (Figura 4).
rpt.3502.fs2104011-figura4.png
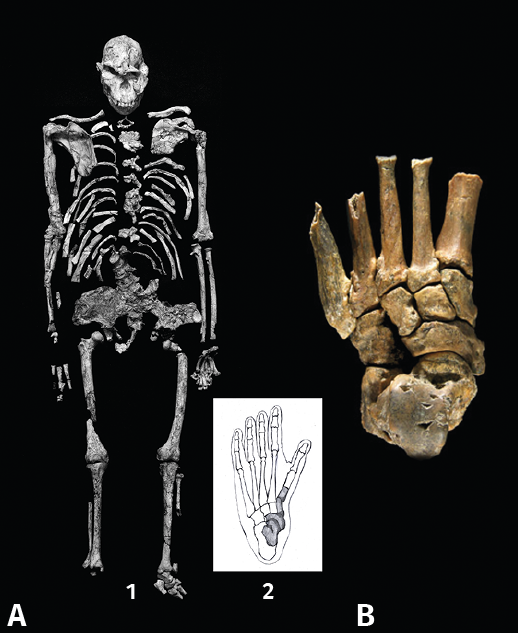
Figura 4. Composición que demuestra que la evolución del pie ha sido en mosaico. A (1 y 2): pie de A. prometheus, “Little foot”, de 3,67 MA; B: pie de OH 8 de Homo habilis de 1,8 MA. La inclinación de la primera articulación tarsometatarsiana tiene un eje muy parecido.
Poco antes de los 4 MA aparecen los Australopithecidos, de los cuales la especie más conocida, en lo que respecta al pie, es el Australopithecus afarensis, en este caso de A.L. 288 – 1 “Lucy”(15). En este estudio se llega a la conclusión de que la 1.ª ATMT en este individuo era ligeramente convexa, a diferencia de otros individuos de la misma especie (A.L.333.28 y A.L.333.79), en los cuales la morfología es ligeramente distinta principalmente en la distribución de las zonas de convexidad articular. Estas y otras diferencias en la morfología del pie en una misma especie demuestran que la adaptación a la marcha bípeda fue discontinua (en “mosaico”) y no tanto una función lineal en el tiempo(16). Es “crucial” para determinar la evolución de estos homínidos la valoración de su postura predominante, así como el tipo de locomoción. Estas características se analizan a través de la aparición de determinados cambios morfológicos en su anatomía. Entre estos cambios, la zona de predilección es el pie(17,18) (Figura 5).
rpt.3502.fs2104011-figura5.png
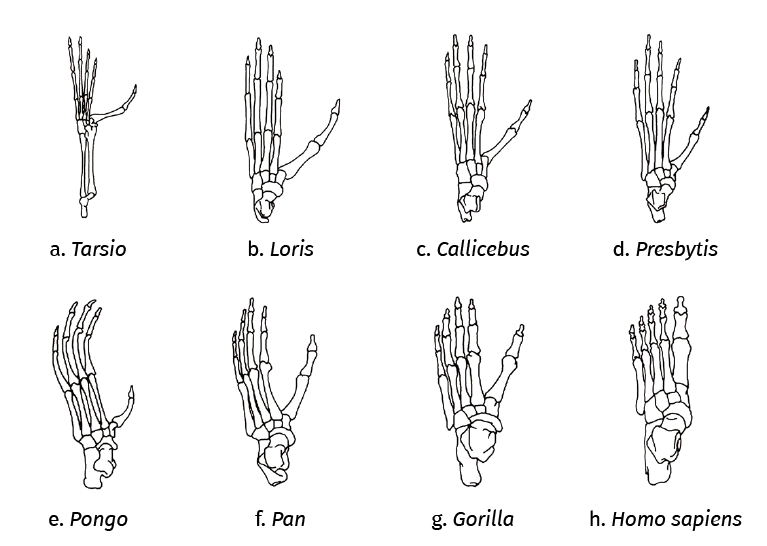
Figura 5. Visión dorsal comparativa del pie de algunos primates. En el dibujo puede verse el grado de divergencia del hallux a partir de la primera articulación tarsometatarsiana. a: Tarsio (35 MA); b: Loris (23 MA) – Prosimii; c: Callicebus (20 MA); d: Presbytis (15 MA) – Simios; e: Pongo (10-8 MA); f: Pan (10-8 MA); g: Gorilla (10-8 MA) – Pongidae; h: Homo sapiens (300.000 a). Modificado de Morton, 1924(17). Cronología adaptada de Fleajgle, 1988(18).
Un exhaustivo estudio antropológico de los restos fósiles de A. afarensis (los más numerosos) ha determinado que la 1.ª ATMT es ligeramente convexa, pero lo más importante es que su morfología es la denominada kidney-shaped, es decir, en forma arriñonada, muy similar a la que presenta el hombre actual. Pero, a diferencia de la del hombre moderno, esta articulación muestra en su cara plantar un tubérculo en el que se inserta el tendón del peroneous longus, circunstancia que determina una capacidad, aunque menor, de aprehensión de este pie(19).
El primer Homo propiamente dicho fue el Homo habilis del Pleistoceno (2,3 MA) de Sudáfrica y África del Este. El pie más estudiado es el del individuo OH 8 de Olduvai. Aunque se trata de un individuo juvenil, las características morfológicas de su pie muestran ya caracteres plesiomórficos o regresivos. En él se conjugan rasgos compatibles con vida arbórea con otros típicos de la marcha bípeda. Una de las principales características será la elongación del mediotarso para favorecer el brazo de palanca, principalmente durante la segunda fase de la marcha bípeda(20).
La 1.ª ATMT es relativamente plana en comparación con sus antecesores, lo que demuestra un primer radio aducido.
Entre los 2,0 y los 1,8 MA, en África y Europa, hace su aparición en el escenario el Homo erectus; las características del pie cambian de una forma bastante radical y se centran en la absorción del peso del cuerpo. Las articulaciones del mediotarso y del complejo subtalar, como consecuencia, pasan a ser articulaciones de amortiguación, perdiendo por ello gran parte de su capacidad móvil intrínseca.
La morfología de la 1.ª ATMT pierde cualquier tipo de funcionalidad móvil intrínseca, derivando hacia una articulación cóncavo-convexa de alto radio, es decir, con una curva muy plana. Dicha característica se incrementa con la aparición del Homo sapiens hace algo más de 300.000 años en el norte de África. Su pie muestra la morfología moderna, con la presencia de los arcos medial y transverso totalmente desarrollados. No obstante, aun en este nivel tan próximo a nosotros, algunas especies como el Homo neanderthalensis muestran caracteres atávicos o regresivos, por lo que durante un tiempo se los denominó “pie ancestral”.
Como conclusión, puede deducirse que el CM y la 1.ª ATMT se han ido modificando a través de los tiempos para adaptarse a los cambios medioambientales y ser lo más funcionales posible. Los primitivos primates tenían una 1.ª ATMT plana, semejante a la que tenemos nosotros, posteriormente el hallux fue progresivamente haciéndose divergente y aprehensor, adquiriendo una forma cóncavo-convexa, para poder desplazarse por los árboles (braquiación). Una vez se consigue la bipedestación y la postura erecta, el hallux pierde la abducción o separación y la articulación de nuevo se hace plana. Esta evolución de la 1.ª ATMT no ha sido pues en forma lineal, sino en mosaico.
Figuras
Figura 1. A: reptil mamiferoide avanzado (Aulacoephalodon peavoti/therapsido); B: detalle del mediotarso de un reptil-mamiferoide avanzado (Bauria cynops/Cynodontia). El radio de la derecha es el primer radio; los 3 huesos que coinciden con el 1.º, 2.º y 3.er radios son los 3 cuneiformes.
Figura 2. Prosimio. A: Nycticebus concuang; B: gran separación entre el primer radio y los radios laterales.
Figura 3. Simio antropoide. A: Cercopithecus aethiops; B: obsérvese la oblicuidad de la primera articulación tarsometatarsiana.
Figura 4. Composición que demuestra que la evolución del pie ha sido en mosaico. A (1 y 2): pie de A. prometheus, “Little foot”, de 3,67 MA; B: pie de OH 8 de Homo habilis de 1,8 MA. La inclinación de la primera articulación tarsometatarsiana tiene un eje muy parecido.
Figura 5. Visión dorsal comparativa del pie de algunos primates. En el dibujo puede verse el grado de divergencia del hallux a partir de la primera articulación tarsometatarsiana. a: Tarsio (35 MA); b: Loris (23 MA) – Prosimii; c: Callicebus (20 MA); d: Presbytis (15 MA) – Simios; e: Pongo (10-8 MA); f: Pan (10-8 MA); g: Gorilla (10-8 MA) – Pongidae; h: Homo sapiens (300.000 a). Modificado de Morton, 1924(17). Cronología adaptada de Fleajgle, 1988(18).

Información del artículo
Cita bibliográfica
Responsabilidades éticas
Protección de personas y animales. Los autores declaran que para esta investigación no se han realizado experimentos en seres humanos ni en animales.
Confidencialidad de los datos. Los autores declaran que han seguido los protocolos de su centro de trabajo sobre la publicación de datos de pacientes.
Derecho a la privacidad y consentimiento informado. Los autores declaran que en este artículo no aparecen datos de pacientes.
Financiación. Los autores declaran que este trabajo no ha sido financiado.
Conflicto de intereses. Los autores declaran no tener ningún conflicto de intereses.
Descargar artículo:
Licencia:
Este contenido es de acceso abierto (Open-Access) y se ha distribuido bajo los términos de la licencia Creative Commons CC BY-NC-ND (Reconocimiento-NoComercial-SinObraDerivada 4.0 Internacional) que permite usar, distribuir y reproducir en cualquier medio siempre que se citen a los autores y no se utilice para fines comerciales ni para hacer obras derivadas.
Comparte este contenido
En esta edición
- Hacia una nueva normalidad
- A veces sucede al revés, el bosque no nos deja ver el árbol
- Evolución de la primera articulación tarsometatarsiana. Un viaje de ida y vuelta
- Anatomía de la primera articulación tarsometatarsiana
- Biomecánica de la primera articulación tarsometatarsiana
- Afectación de la primera articulación tarsometatarsiana en las malformaciones del neonato: pie equinovaro, metatarso aducto, pie en serpentín
- ¿Existe una relación entre la primera articulación tarsometatarsiana y el <em>hallux valgus</em>?
- La primera articulación tarsometatarsiana en el pie plano y el pie cavo
- Artrosis de la primera articulación tarsometatarsiana primaria y postraumática secundaria a fractura-luxación de Lisfranc
- Osteotomías distales metatarsales mínimamente invasivas guiadas por ecografía (US-DMMO): estudio de concordancia ecográfica inter- e intraobservador
- Lesiones óseas en pacientes erróneamente diagnosticados de esguince de tobillo. Serie de casos y factores a tener en cuenta
- Técnica de extracción de un clavo de artrodesis tibiotalocalcánea roto. Serie de casos
- Síndrome del túnel del tarso posterior. Tratamiento y revisión. A propósito de un caso
- Revista de revistas
Más en PUBMED
Más en Google Scholar
Más en ORCID

Revista del pie y tobillo está distribuida bajo una licencia de Creative Commons Reconocimiento-NoComercial-SinObraDerivada 4.0 Internacional.